SINGLE
NUCLEOTIDE POLYMORPHISMS OF THE HEAT SHOCK PROTEIN 90 GENE IN VARICOCELE-ASSOCIATED
INFERTILITY
(
Download pdf )
PERICLES A. HASSUN FILHO, AGNALDO P. CEDENHO, SAMIRA B. LIMA, VALDEMAR ORTIZ, MIGUEL SROUGI
Department of Surgery, Division of Urology, Federal University of Sao Paulo, Sao Paulo, Brazil
ABSTRACT
Purpose:
Varicoceles are associated with impaired testicular function and male
infertility, but the molecular mechanisms by which fertility is affected
have not been satisfactorily explained. Spermatogenesis might be affected
by increased scrotal temperature, such as that caused by varicocele. HSP90
is a molecular chaperone expressed in germ cells and is related to spermatogenesis,
motility, and both heat and oxidative stress. Possible correlations between
coding single region nucleotide polymorphisms (cSNPs) in the HSP90 gene
in patients with varicocele associated with infertility were analyzed,
and polymorphisms in these exons were characterized through DNA sequencing.
Materials and Methods: PCR-SSCP and DNA
sequencing were used to search for mutations in 18 infertile patients
with varicocele, 11 patients with idiopathic infertility and 12 fertile
men. DNA was extracted from leucocytes for PCR amplification and SSCP
analysis. DNA from samples with an altered band pattern in the SSCP was
then sequenced to search for polymorphisms.
Results: Three silent polymorphisms that
do not lead to amino acid substitutions were identified.
Conclusion: Mutations in the HSP90 gene
do not appear to be a common cause of male factor infertility. The low
incidence of gene variation, or SNPs, in infertile men demonstrates that
this gene is highly conserved and thus confirms its key role in spermatogenesis
and response to heat stress.
Key
words: varicocele; heat stress; Heat-Shock Protein 90; spermatozoa;
infertility, genetic polymorphism
Int Braz J Urol. 2005; 31: 236-44
INTRODUCTION
The
male factor is responsible for 51.2% of conjugal infertility and 39% of
these men present an abnormal semen analysis for idiopathic reasons. On
the other hand, varicocele is shown to be the second most common cause
of male infertility leading ultimately to lower testicular volumes and
important seminal alteration (1,2). Despite all this evidence there is
controversy as to the mechanisms by which varicocele hinders spermatogenesis
and affects the fertilizing potential in men (3).
Many clinical observations suggest a link
between testicular hyperthermia and reduction of spermatogenesis by apoptosis,
such as those caused by varicocele (4). However, the molecular mechanisms
of gene expression affected by higher temperatures and the events that
activate cell death under this condition remain unknown; these factors
could be identified by specific molecular/genetic markers (5). The human
genome variability may validate key evidence in understanding individual
susceptibility to complex diseases. A molecular genesis for seminal alterations
has been found in patients with varicocele (6), which provides new information
about this disease, as well as new diagnostic possibilities.
Although some genes are expressed in both
germ and somatic cells, others are exclusively expressed in germ cells.
Any disruption to the regular expression of these genes may lead to abnormal
spermatogenesis (7). The highly preserved “Heat Shock Protein”
(HSP) family of genes (8) is extremely important, since these genes act
as molecular chaperones in a wide variety of cellular processes. HSP70
and HSP90 are expressed in germ, Sertoli and Leydig cells during the neo-natal
period, and in spermatocytes and spermatids after the onset of puberty;
HSP90 also expresses in spermatogonias (9). These proteins reestablish
a homeostatic mechanism and equilibrium between protein synthesis and
degradation in the cells (10). This has been shown in the heat-induced
differential expression of HSP70 and HSP90 in varicocele (11,12) and in
semen cryo-preservation (13).
As well as being responsive to stress, HSP90
is a highly preserved protein that is highly abundant – between
1 and 3% of the total amount of protein found in most cells is HSP90 (14).
This molecular chaperone plays an important role in spermatogenesis (15),
and reduced HSP90 function in transheterozygote Drosophila sp correlates
with an infertility phenotype (16). In a study conducted by Huang et al.
(2000) with boar semen, the authors concluded that through direct action
in these cells, HSP90 is responsible for a reduction of sperm motility
(17). HSP90 is also associated with protecting the cells against oxidation
damage (18).
Yue et al. (1999) suggest that a small change
in HSP90 function, such as point mutations, could lead to infertility
(16). It has also been reported that HSP90’s role in protein folding
and maturation could buffer the damaging effects of mutations (19).
In order to define the mechanisms underlying
HSP90 function, Rebbe et al. (1989) determined its genomic structure,
which is built of 12 exons and 11 introns (20), and as a result screening
for mutations in infertile men with varicocele has become possible.
This study set out to analyze possible correlations
between coding single region nucleotide polymorphisms (cSNPs) in the HSP90
gene using PCR-SSCP (polymerase chain reaction – single strand conformation
polymorphism) in patients with varicocele associated with azoospermia
or severe oligozoospermia, and to characterize possible mutations in these
exons through DNA sequencing.
MATERIALS AND METHODS
A
prospective study was employed involving three groups of patients: Group
A, made up of 18 infertile patients with grade II or III varicocele (14
bilateral and 4 unilateral varicoceles), of which 12 presented severe
oligozoospermia (< 5x106 sperm/mL) and 6 presented azoospermia; Group
B, made up of 11 patients with idiopathic infertility, of which 6 presented
severe oligoasthenozoospermia and 5 presented azoospermia; and Group C,
made up of 12 patients with proven fertility and without varicocele. Testicular
volume was assessed in all patients through the use of a Prader orchidometer,
and blood samples were collected for FSH, LH and serum testosterone dosage
and for genetic analyses.
Each patient was fully informed of the objectives
of this study and those that agreed with the consent form were assigned
to the study. All protocols were first approved by the Research Ethics
Committee.
Semen Analysis - Semen was collected twice
for each patient and the samples were analyzed according to the World
Health Organization guidelines (2) and sperm morphology was evaluated
using strict criteria according to Kruger et al. (1996) (21).
Hormonal Analysis - A blood plasma aliquot
maintained at -20oC was used for luteinizing hormone (LH) and follicle
stimulating hormone (FSH) dosage by enzyme immunoassay using the IMÒxsystem
kit from Abbott Laboratories, according to the manufacturer’s protocol.
Results were expressed in mIU hormone/mL plasma. Reference values for
LH and FSH are respectively 2.0 to 12.0mIU/mL and 1.0 to 12.0mIU/mL. Another
blood plasma aliquot, also maintained at -20oC, was tested by radioimmunoassay
in solid phase to determine testosterone levels using the Coat-A-Count
Total Testosterone (DPC) kit. Results were expressed in ng testosterone/dL
plasma. Intra-assay variation was determined at 1.8% and inter-assay at
2.1%. Reference values for 20 to 49 year old men are from 262 to 1.836ng/dL.
DNA analysis - All infertile patients were
submitted to the Y chromosome microdeletion scan using multiplex PCR for
regions AZFa, AZFb, AZFc, and AZFd. 18 pairs of oligonucleotides that
are homologous to the following STS (sequence tagged sites) were used:
sY81, sY182, sY121, sYPR3, sY124 sY127, sY128, sY130, sY133, sY145, sY153,
sY152, sY242, sY239, sY208, sY254, sY255, and sY157. Patients showing
Y chromosome microdeletions were submitted for further analysis of chromosome
abnormalities using peripheral lymphocyte culture and G band staining.
Based on the previously described DNA sequences
for the HSP90 gene, GeneBank access J04988, primer pairs for PCR were
designed from the 5' and 3' flanking regions for each of the 11 coding
HSP90 gene exons (Table-1).

Purified DNA samples from leukocytes were
used for the PCR reactions. Each reaction was prepared to a final volume
of 50µL, containing: DNA (100ng), 1.25mM of each deoxyribonucleotide
triphosphate (dNTP), Taq DNA polymerase (2IU), 10x concentrated buffer
for Taq DNA polymerase, bovine serum albumin (BSA), 2.5mM magnesium chloride
(MgCl2), and ultra-pure water q.s.p. The temperature cycles used in each
reaction were based on previous annealing temperature analyses for each
pair of oligonucleotides. Negative controls containing all the reagents
except DNA were used in all the experiments to monitor for possible DNA
contamination.
The PCR amplified DNA fragments for all
11 exons of the HSP90 gene were precipitated through addition of sodium
acetate pH 6.0 to a final concentration of 0.3M and absolute ethanol and
further incubation at -20oC for 30 minutes. After this period the tubes
were centrifuged at 13,000 rpm and the sediment was re-suspended in 20
µL of TE buffer (10mM Tris pH 7.4 and 1mM EDTA). A CD buffer (05%
formamide, 0.05% xylene cyanol, 0.05% bromophenol blue and 20mM EDTA)
was added to the samples, which were heated for 10 minutes prior to electrophoresis.
The PCR product was separated into a 12%
polyacrilamide horizontal gel, GeneGel Excel 12.5/24 (Amersham Biosciences).
The gel was run in a GenePhor electrophoresis chamber (Amersham Biosciences)
under the following conditions: 10-15oC, 110V, 16mA, 6W for 20 minutes;
10-15oC, 600V, 42mA, 16W for 120 minutes.
DNA was observed using the PlusOne DNA Silver
Staining kit (Amersham Biosciences) in the GeneStain Automated Gel Stainer
automatic unit (Amersham Biosciences).
PCR products were cloned into TOPO TA Cloning
Kit vectors (Invitrogen), according to the manufacturer’s protocol.
Sequencing reactions were performed using the Dye Primer Cycle Sequencing
Ready Reaction reagent kit (Perkin Elmer) according to manufacturer’s
instructions. Electrophoresis was performed in the automatic ABI 377 equipment
(Perkin Elmer). All PCR products were sequenced in both ways (forward
and reverse), and sequencing was repeated once.
The sequences were then compared to those
of the HSP90 gene found in the GeneBank database.
RESULTS
Mutation
analyses were performed in the 11 coding HSP90 gene exons. Since the first
exon does not codify for a protein, it wasn’t evaluated. The others
were analyzed (Table-1) according to the previously described methodology.
Seven altered conformation patterns were detected with the SSCP analysis.
Figure-1 shows a polyacrilamide gel representing exon 10 of the HSP90
gene.
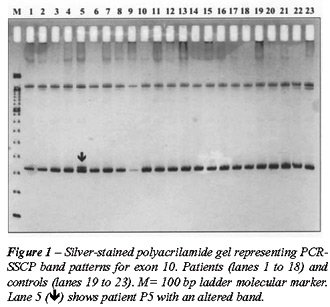
DNA from samples showing an abnormal band
pattern on the SSCP analysis were sequenced in order to search for mutations.
Three SNPs were identified: a 3774 C>T polymorphism in exon 4 of patient
P11; a 6192G>A polymorphism in exon 10 of patient P5; and a 7602G>A
polymorphism in exon 12 of patient P8 (Figures-2, 3 and 4).
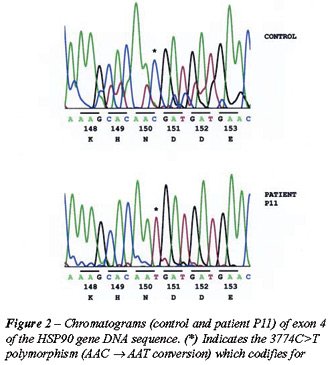
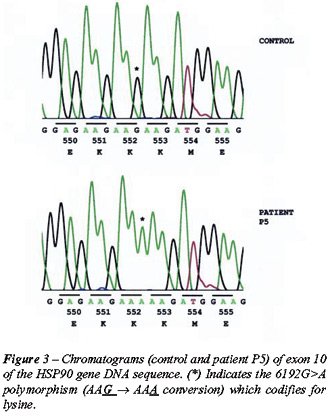
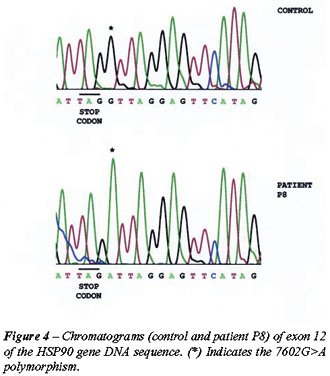
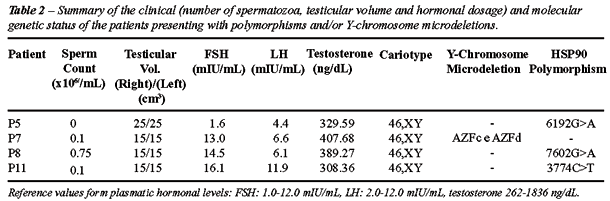
Table-2 summarizes the clinical and hormonal
status of the 3 patients showing polymorphisms for the HSP90 gene, and
the patient with Y chromosome microdeletions. Patient P7, who presented
oligoasthenozoospermia, did not show any polymorphisms in the HSP90 gene
but was tested positive for microdeletions in the AZFc and AZFd Y chromosome
regions, and a raised FSH concentration (13.0 mIU/mL). Patient P5 presented
azoospermia and tested normal for testicular volume and hormonal levels.
Patients P8 and P11 presented varicocele and were diagnosed with severe
oligozoospermia, asthenozoospermia and raised FSH levels (14.5 and 16.1
mIU/mL respectively).
COMMENTS
Varicocele
is one of the most frequent causes of male infertility, although the pathogenic
mechanisms by which it leads to changes in spermatogenesis are not clear.
Some of these mechanisms may possess a known cause, such as Y-chromosome
microdeletions. In this study, the frequency of Y-chromosome microdeletions
was assessed. Of the 29 patients investigated, 1 (patient P7) with severe
oligozoospermia presented microdeletions in Y chromosome regions AZFc
and AZFd, a frequency of 3.45%, which is within the classically described
range of 3-20% (22).
As well, 18 patients with azoospermia or
severe oligozoospermia associated with varicocele and 11 patients with
idiopathic infertility were screened for DNA polymorphisms in the HSP90
gene. Mutations in this gene could lead to oligoasthenoteratozoospermia
in men similar to that shown by patients with Y chromosome microdeletions.
Of all the samples analyzed, only three alterations were found, which
characterizes the high conservation rate of this protein (8). Patient
P11 possessed a 3774C>T polymorphism in codon 150, exon 4, which codifies
for asparagine (Figure-2); Patient P5 possessed a 6192G>A polymorphism
in codon 552, exon 10, which codifies for lysine (Figure-3); and Patient
P8 possessed a 7602G>A polymorphism in the non-coding region (3’UTR)
of exon 12, one base after the UAG stop-codon (Figure-4). Although these
synonymic SNPs do not alter the protein sequence, they could lead to differences
in messenger RNA (mRNA) structure and stability and affect the amount
of protein produced (23).
The three SNPs found are transitions, substitutions
from a purine to a purine or a pyrimidine to a pyrimidine. These alterations
are sometimes originated in base incorporation errors during DNA replication.
If these mutations occur in germ cells, they may be transmitted to future
generations and fixate in the population. This is particularly important
when genetic causes for idiopathic infertility are sought, especially
in candidates for intracytoplasmic sperm injection (ICSI).
Although there is not yet any treatment
to improve fertility in patients with genetic alterations, these studies
have been important in male infertility diagnosis. Molecular diagnosis
allows the clinician to offer adequate counseling to the patient who wishes
to be a biological father, since ICSI bypasses natural selection mechanisms.
Therefore, as in other areas of andrology, more basic research is needed
to study varicocele etiology, especially to identify the infertility associated
traits.
CONCLUSION
Mutations of the HSP90 gene do not seem to be a common cause of male factor infertility. The low incidence of gene variation or SNPs in infertile men demonstrates the high conservation rate of this gene and thus confirms its key role in spermatogenesis and response to stress.
REFERENCES
- World Health Organization WHO: Towards more objectivity in diagnosis and management of male infertility. Results of a World Health Organization multi-centre study. Int J Androl. 1987; Suppl 7, 10: 1-35.
- World Health Organization WHO: Laboratory manual for the examination of human semen and sperm-cervical mucus interaction. 4th ed. Cambridge University Press, 1999.
- Schlesinger MH, Wilets IF, Nagler HM: Treatment outcome after varicocelectomy. A critical analysis. Urol Clin North Am. 1994; 21: 517-29.
- Mieusset R, Bujan L: Testicular heating and its possible contributions to male infertility. Int J Androl. 1995; 18: 169-84.
- Nakai A, Suzuki M, Tanabe M: Arrest of spermatogenesis in mice expressing an active heat shock transcription factor 1. EMBO J. 2000; 19: 1545-54.
- Marmar JL: The pathophysiology of varicoceles in the light of current molecular and genetic information. Hum Reprod Update. 2001; 7: 461-72.
- Feng HL, Sandlow JI, Sparks AE: Decreased expression of heat shock protein HSP70-2 is associated with the pathogenesis of male infertility. Fertil Steril. 2001; 76: 1136-9.
- Neuer A, Spandorfer SD, Giraldo P, Dieterle S, Rosenwaks Z, Witkin SS: The role of heat shock proteins in reproduction. Hum Reprod Update. 2000; 6: 149-59.
- Ogi S, Tanji N, Iseda T, Yokoyama M: Expression of heat shock proteins in developing and degenerating rat testes. Arch Androl. 1999; 43: 163-71.
- Shi Y, Mosser DD, Morimoto RI: Molecular chaperones as HSF1-specific transcriptional repressors. Genes Dev. 1998; 12: 654-66.
- Brackin P, Marmar JL, Camden NJ, Napolitano B, Millan C, Benoff S: Identification of Heat Shock Protein 70-1 (HSP70-1) in testis tissue of men with varicoceles. J Urol. 2003; 169: 415.
- Lima SB, Hassun PA, Cenedese MA, Cedenho AP, Srougi M: Expression of the HSP70-2 gene in sperm cells of adolescents with varicocele. J Urol. 2003; 169: 452.
- Cao WL, Wang YX, Xiang ZQ, Li Z: Cryopreservation-induced decrease in heat-shock protein 90 in human spermatozoa and its mechanism. Asian J Androl. 2003; 5: 43-6.
- Lai BT, Chin NW, Stanek AE, Keh W, Lanks KW: Quantitation of and intracellular localization of the 85-k heat-shock protein by using monoclonal and polyclonal antibodies. Mol Cell Biol. 1984; 4: 2802-10.
- Gruppi CM, Zakeri ZF, Wolgemuth DJ: Stage and lineage-regulated expression of two hsp90 transcripts during mouse germ cell differentiation and embryogenesis. Mol Reprod Dev. 1991; 28: 209-17.
- Yue L, Karr TL, Nathan DF, Swift H, Srinivasan S, Lindquist S: Genetic analysis of viable Hsp90 alleles reveals a critical role in Drosophila spermatogenesis. Genetics. 1999; 151: 1065-79.
- Huang SY, Kuo YH, Tsou HL, Lee YP, King YT, Huang HC, et al.: The decline of porcine sperm motility by geldanamycin, a specific inhibitor of heat-shock protein 90 (HSP90). Theriogenology. 2000; 53: 1177-84.
- Conconi M, Szweda LI, Levine RL, Stadtman ER, Friguet B: Age-related decline of rat liver multi-catalytic proteinase activity and protection from oxidative inactivation by heat-shock protein 90. Arch Biochem Biophys. 1996; 331: 232-40.
- Queitsch C, Sangster TA, Lindquist S: HSP90 as a capacitator of phenotypic variation. Nature. 2002; 417: 618-24.
- Rebbe NF, Hickman WS, Ley TJ, Stafford DW, Hickman S: Nucleotide sequence and regulation of a human 90-kDa heat shock protein gene. J Biol Chem. 1989; 264: 15006-11.
- Kruger TF, Lacquet FA, Sarmiento CA, Menkveld R, Ozgur K, Lombard, CJ, et al.: A prospective study on the predictive value of normal sperm morphology as evaluated by computer (IVOS). Fertil Steril. 1996;66: 285-91.
- Maurer B, Gromoll J, Simoni M, Nieschlag E: Prevalence of Y chromosome microdeletions in infertile men who consulted a tertiary care medical center: the Munster experience. Andrologia. 2001; 33: 27-33.
- Kwok PY, Gu Z: Single nucleotide polymorphism libraries: why and how are we building them? Mol Med Today. 1999; 5: 538-43.
________________________
Received: February 24, 2005
Accepted after revision: April 30, 2005
_______________________
Correspondence address:
Dr. Péricles Assad Hassun Filho
R. Leandro Dupret, 204 / 43
São Paulo, SP, 04025-010, Brazil
Fax: + 55 11 5575-9077
E-mail: pericles.hassun@gmail.com
EDITORIAL COMMENT
VARICOCELES, HEAT AND HEAT SHOCK PROTEINS - THAT IS THE QUESTION!
The
variable effect of the varicocele on spermatogenesis remains enigmatic.
Why do varicoceles affect spermatogenesis? Why not all patients with varicoceles
have abnormal spermatogenesis? An understanding of the pathophysiology
of the varicocele will allow patients and their physicians to choose the
most appropriate therapy for their infertility and varicocele.
In the current article by Hassun Filho et
al., the molecular basis of varicocele associated abnormal spermatogenesis
is explored.
The authors studied single nucleotide polymorphisms
of the heat shock protein (HSP) 90 gene in men with varicocele associated
infertility. Heat-shock proteins are molecular chaperones, which assist
other proteins in their folding, transport and assembly into complexes,
thus protecting both protein structure and function. These heat shock
proteins are most often induced by physiological stress such as heat (heat
shock protein) and other stresses. Heat shock proteins are classified
according to their molecular weight: HSP25, HSP47, HSP60, HSP70/72, HSP90,
and HSP110 (1). Thus, their expression in the testicle protects the germ
cells from environmental hazards such as heat, radiation, and chemicals
(2). The activation of this genetically programmed response to stress
has been evolutionary conserved (3). Thus, suggesting that mutations affecting
HSP90 could theoretically lead to infertility (4). Therefore, such mutations
could limit the protective effects of heat shock protein in response to
the stress induced by varicoceles.
Some have suggested that HSP dysregulation
in varicocele patients initiates germ cell apoptosis, and, it follows
that coding and noncoding mutations could interfere with the normal protein
kinetics of HSP90 (5). Simsek et al. documented increased apoptosis in
patients with varicoceles (6).
Other attempts have been made to explain
the variable impact of the varicocele on spermatogenesis. Nagler and coworkers
constructed the The Cofactor Hypothesis of the varicocele in 1990 (7).
This hypothesis states that, perhaps, the varicocele is a cofactor, which
allows other gondatoxins to manifest themselves. This work demonstrated
that nicotines’ effect on spermatogenesis was more pronounced when
administered in conjunction with an experimental varicocele. Marmar &
Benoff subsequently proposed a “two hit hypotheses” (8). This
hypothesis states that molecular or genetic defects may be required for
the varicocele to exert its deleterious effects on spermatogenesis. In
the absence of such defects, men with varicoceles may be fertile. It is
this later hypothesis that is studied in the current manuscript.
The current study, unfortunately, does not
allow us to conclude that mutations in the HSP cascade are the underlying
pathophysiologic mechanism of the abnormal spermatogenesis associated
with the varicocele. The assessment of Y chromosome microdeletions although,
as the authors note, is consistent with published data for infertile men
does not relate to the varicocele. If mutations in HSP were identified
in the men with varicoceles and infertility, it would perhaps provide
evidence that disruption in this highly conserved protective mechanism
could explain the variable impact of the varicocele on spermatogenesis.
Nevertheless, there is little doubt the genetic/molecular studies of infertility
will ultimately provide additional understanding. Alas, no differences
in the HSP genes were observed in this small population. Thus, we are,
again left questioning why does a varicocele affect spermatogenesis when
it does and why does not the varicocele always affect spermatogenesis.
REFERENCES
- Morimoto RI, Tissieres A, Georgopoulos C: Progress and Perspectives on the Biology of Heat Shock Proteins and Molecular Chaperones. Cold Spring Harbor, Cold Spring Harbor Press, 1994.
- Legare C, Thabet M, Sullivan R: Expression of heat shock protein 70 in normal and cryptorchid human excurrent duct. Mol Hum Reprod. 2004; 10: 197-202.
- Feder ME, Hofmann GE: Heat-shock proteins, molecular chaperones, and the stress response: Evolutionary and ecological physiology. Annu Rev Physiol. 1999; 61: 243-82.
- Yue L, Karr TL, Nathan DF, Swift H, Srinivasan S, Lindquist S: Genetic analysis of viable Hsp90 alleles reveals a critical role in Drosophila spermatogenesis. Genetics. 1999; 151: 1065-79.
- Lue YH, Hikim AP, Swerdloff RS, Im P, Taing KS, Bui T, et al.: Single exposure to heat induces stage-specific germ cell apoptosis in rats: role of intratesticular testosterone on stage specificity. Endocrinology. 1999: 140: 1709-17.
- Simsek F, Turkeri L, Cevik I, Bircan K, Akdas A: Role of apoptosis in testicular damage caused by varicocele. Arc Esp Urol. 1998; 51: 947-50.
- Peng BC, Tomashefsky P, Nagler HM: The cofactor effect: varicocele and infertility. Fertil Steril. 1990; 54: 143-8.
- Marmar J, Benoff S: New scientific information related to varicoceles. J Urol. 2003; 170 (6 pt 1): 2371-3.
Dr.
Harris M. Nagler
Beth Israel Medical Center and
Albert Einstein College of Medicine
New York, New York, USA