EFFECTS OF MALNUTRITION IN THE TESTIS
ELAINE C. MOTA, ALBA M. SANTOS, FABIANE P. TOSTE, FRANCISCO J.B. SAMPAIO, CRISTIANE F. RAMOS
Laboratory of Molecular Biology, Urogenital Research Unit, State University of Rio de Janeiro (UERJ), Rio de Janeiro, RJ, Brazil
ABSTRACT
Introduction:
Androgens are important to spermatogenesis in the testes, to sperm maturation
in the epididymis and to development and function of male accessory glands.
The goal of this study is to investigate the effect of mother’s malnutrition
in the androgen receptor expression of post-weaning rat testis.
Material and Methods: At delivery, Wistar
rats were assigned to one of the following groups: control (C) = diet
with 23% of protein; protein-restricted (PR) = diet with 8% of protein;
energy-restricted (ER) = diet with 23% of protein in restricted quantities,
according to the ingestion of PR group. At the end of lactation period,
the animals were sacrificed, the testes excised, weighted and the androgen
receptor determined by Western blotting technique.
Results: Testicular weight was significantly
lower (p < 0.001) in both groups at the end of lactation, when compared
to controls (64.7% in the PR and 60.1% in the ER group). Both PR and ER
groups had a significant (p < 0.001) increase in the relative quantity
of androgen receptor, but this increment was higher in the PR group.
Conclusion: The observed increase in the
relative quantity of androgen receptor in response to the maternal malnutrition
can be consequent to alterations in the serum concentrations of gonadotropins,
estrogen and androgen, since the androgen receptors are regulated by these
factors.
Key words:
testis; malnutrition; rats; androgen receptor; Western blotting
Braz J Urol, 27: 500-506, 2001
INTRODUÇÃO
Os
hormônios androgênios são importantes para a manutenção
da espermatogênese no testículo, para a maturação
do espermatozóide no epidídimo e para o desenvolvimento
e funções dos órgãos sexuais acessórios
masculinos (1).
Através
de técnicas de imunocitoquímica, o receptor de androgênio
(AR) foi localizado em tecidos do sistema reprodutor de homens e ratos
de ambos os sexos (2,3). O receptor de androgênio é uma proteína
com peso molecular entre 100 e 110 kd (4), já tendo sido clonada
(5,6).
Os
níveis de AR são regulados por androgênio nas células
de Sertoli de animais adultos (7,8), enquanto em animais imaturos o hormônio
folículo estimulante (FSH) parece ser o principal regulador (9,10).
Shan et al. (11) demonstraram ainda que nas células de Leydig ocorre
o inverso, os níveis de AR são estimulados por androgênios
nos animais imaturos, não tendo efeito nos animais adultos.
Cardone
et al. (12) utilizando cultura de células testiculares demonstraram
que tanto estrogênios como androgênios podem autoregular a
expressão dos RNA mensageiros dos seus próprios receptores,
e ainda que os estrogênios podem diminuir a expressão dos
RNA mensageiros nos receptores de androgênio. Foi demonstrado que
o androgênio circulante, independente do conteúdo luminal,
é responsável pela regulação da expressão
dos receptores de androgênio (13).
A
restrição alimentar pode inibir tanto a manutenção
como o início da capacidade reprodutiva (14,15). Camundongos adultos
submetidos à restrição alimentar de 30% por até
8 semanas apresentam redução de 42% na concentração
de testosterona sérica, redução de 27% na massa testicular
e um aumento na apoptose, que pode ser responsável pela regressão
testicular que ocorre em resposta à restrição alimentar
(16).
Não
encontramos na literatura estudos sobre os efeitos da desnutrição
materna no período de lactação sobre os tecidos do
sistema urogenital de ratos. Assim, o presente estudo tem como objetivo
investigar a expressão dos receptores de androgênio nos testículos
de filhotes de ratas lactantes desnutridas.
MATERIAL E MÉTODOS
Ratas
Wistar fêmeas, nulíparas, foram mantidas em biotério
com temperatura estável (25 ± 10C) e ciclo claro-escuro
(7:00h - 19:00h) controlados. Foram utilizados os princípios descritos
em “The Guide for the Care and Use of Laboratory Animals” para
os cuidados no uso e manuseio dos animais (17).
Aos
3 meses de idade estes animais foram acasalados na proporção
de 2 fêmeas para 1 macho, recebendo ração comercial
(23% de proteína) até o nascimento dos filhotes, quando
então foram divididas em 3 grupos de tratamento:
1)- Controle (C): com livre acesso a água e a dieta normal (ração
comercial com 23% de proteína).
2)- Restrição protéica (RP): com livre acesso a água
e a dieta hipoprotéica (8% de proteína).
3)- Restrição calórica (RC): com livre acesso a água,
porém submetido a dieta normal (ração comercial com
23% de proteína) restrita às mesmas quantidades ingeridas
no dia anterior pelo grupo em dieta hipoprotéica.
O
grupo de animais em restrição calórica, pareados
pela quantidade de ração ingerida pelo grupo em dieta hipoprotéica,
foi estudado a fim de diferenciar os efeitos causados pela carência
exclusiva de proteína daqueles causados pela restrição
calórica, uma vez que já foi demonstrado que ratas lactantes
submetidas à restrição protéica ingerem uma
quantidade menor de ração em relação às
ratas controles (18,19).
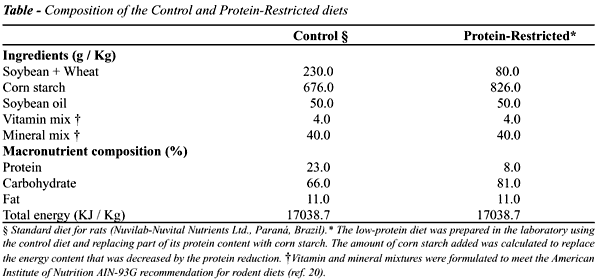
A
dieta hipoprotéica foi preparada manualmente em nosso laboratório
e sua composição é mostrada na Table. A fonte protéica
(8%) desta dieta foi a ração comercial macerada (Labina
- Purina Nutrimentos Ltda.) e as calorias foram compensadas pelo acréscimo
de amido de milho a fim de se obter uma dieta hipoprotéica e isocalórica.
As vitaminas e os minerais foram suplementados de maneira a se obter a
mesma composição da ração comercial que é
baseada nas recomendações do National Research Council e
do National Institute of Health, USA (20).
A
dieta hipoprotéica foi administrada a partir do dia do nascimento
dos filhotes quando a ninhada foi ajustada em número de 6. Este
número de filhotes foi escolhido porque segundo Fishbeck &
Rasmussen parece ser este o número de animais que confere o maior
potencial lactotrófico (21). Provavelmente, o tamanho da ninhada
influencia muito mais na quantidade de leite produzida do que em sua composição
(22).
Aos
21 dias de lactação os animais foram anestesiados com tiopental
(0.1 ml/100g peso corporal), os testículos foram excisados, pesados
e armazenados a –700C para posterior determinação dos
receptores de androgênio pela técnica de Western blotting.
Os
testículos foram homogeneizados em tampão de homogeneização
(50mM TRIS pH 7.4, 1.5mM EDTA, 50mM NaCl, glicerol 10%, 5mM DTT, 10mg/ml
de leupeptina e 30ml/g tecido de PMSF) em um volume de 250ml. O homogenato
foi então centrifugado a 100.000 Xg, por 2h, a 40C. O sobrenadante
foi coletado e armazenado a –200C. A quantidade de proteína
total do sobrenadante foi determinada utilizando-se o método de
Bradford (23).
As
proteínas foram separadas em gel de acrilamida 8% (SDS-PAGE). Foram
adicionados cerca de 60 mg de proteína das amostras em cada slot
do gel. Logo após, as amostras foram transferidas para uma membrana
de nitrocelulose, a qual foi bloqueada com tampão towbin (31mM
TRIS pH 7.4, 3.4 mM KCl, 0.17M NaCl, 0.05mM vermelho de fenol, 0.1% tween
20) contendo 5% de leite desnatado, por 1h, com agitação.
Posteriormente a membrana foi incubada com anticorpo específico
(rabbit polyclonal 200 mg/ml, Santa Cruz proced) em tampão towbin
contendo 0.25% de leite desnatado “overnight”, com agitação,
lavada 3 vezes com tampão towbin, seguindo-se então uma
incubação adicional com o segundo anticorpo (peroxidase-conjugated
donkey IgG, 1:2000), por 1h, com agitação. Após lavagem
da membrana 3 vezes com tampão towbin, a membrana foi incubada
com reagente de detecção ECL (Amersham) por 5 minutos e
exposta num hiperfilme ECL para autoradiografia (Amersham). A densitometria
foi feita utilizando o programa de computador Scion Image.
As
diferenças entre os grupos foram determinadas através de
análise de variância univariada associada a teste de comparação
múltipla entre as médias, teste de Newman-Keuls (24).
RESULTADOS
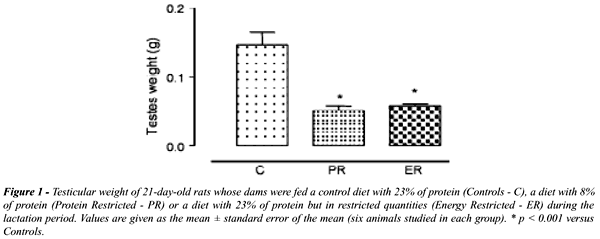
O
peso do testículo dos filhotes de ratas submetidas a restrição
protéica ou calórica na lactação apresentou
redução significativa (p < 0.001) e está representado
na Figure-1. A redução observada no peso do testículo
do grupo Restrição Protéica (RP) é de 64.7%
e do Restrição Calórica (RC) é de 60.1%.
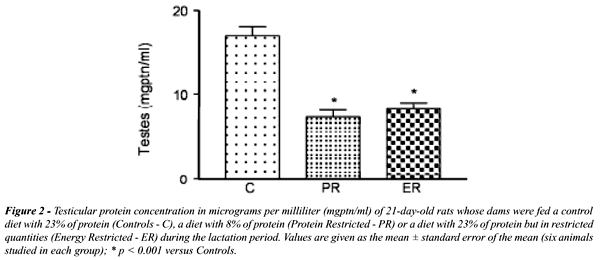
A
concentração de proteína nos testículos dos
filhotes de ratas lactantes desnutridas diminuiu significativamente(Figure-2),
porém quando analisamos por grama de tecido esta diferença
desaparece.
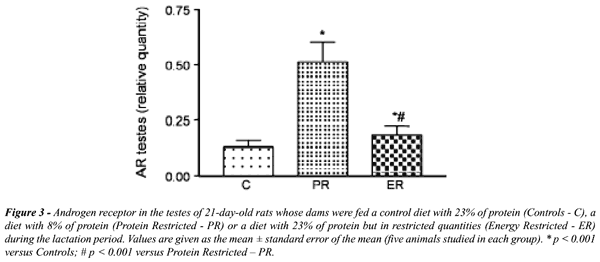
Os
animais cujas mães foram submetidas a ambos os tipos de desnutrição
apresentaram uma quantidade de receptor significativamente maior que os
animais controle, sendo que o aumento no grupo RP foi significativamente
maior que no RC (Figure-3).
DISCUSSÃO
A
desnutrição materna na lactação levou a uma
redução significativa no peso do testículo, o que
está de acordo com Young et al. (16) que relatou diminuição
de 27% no testículo de camundongos adultos desnutridos.
Acreditamos
que a concentração de proteínas totais no testículo
foi reduzida pela desnutrição materna em decorrência
da redução no peso do testículo, já que quando
relacionamos a concentração protéica por grama de
tecido esta alteração não foi mais observada.
A
expressão de receptores de androgênio (AR) nos vários
tecidos ou células varia em resposta ao androgênio. Os níveis
de RNA mensageiro dos AR, em resposta ao androgênio, aumentam no
rim de aves (25) e no músculo liso do pênis de rato (26),
enquanto diminuem no fígado de aves (25) e na próstata ventral
de ratos (7). Ao nível protéico, os AR parecem diminuir
na próstata e vesículas seminais de rato em resposta ao
androgênio.
Zhu
et al. (8) demonstraram que os AR são regulados pelo androgênio
circulante, de maneira diferente nas células de Sertoli, de Leydig
e no epidídimo de ratos adultos. Além disto, alguns autores
sugerem que o FSH e não o androgênio seja o regulador principal
da expressão dos AR em testículos de animais imaturos (9,10).
Cardone
et al. (12) utilizando cultura de células testiculares demonstraram
ainda que o estrogênio pode diminuir a expressão dos RNA
mensageiros dos receptores de androgênio.
Em
macacos adultos, foi demonstrado por Console et al. (28) que a desnutrição
protéica leva a mudanças morfométricas e ultra-estruturais
nos gonadotrofos, com diminuição na densidade volumétrica
e celular e sinais compensatórios de hiperfunção.
Com
base nesses relatos podemos sugerir que a desnutrição materna
pode acarretar alterações nas concentrações
séricas de gonadotrofinas lipofisárias (LH, FSH), androgênios
ou estrogênios na prole, justificando o aumento encontrado nos receptores.
Outros estudos, com dosagem das concentrações séricas
de LH, FSH, androgênios e estrogênios, junto com a determinação
dos AR no testículo estão sendo conduzidos para confirmar
estes achados.
______________________________________
Pesquisa financiada pela Fundação de Amparo
à Pesquisa do Estado do Rio de Janeiro (FAPERJ) e pelo Conselho
Nacional de Desenvolvimento
Científico e Tecnológico (CNPq) Brazil.
REFERÊNCIAS
- Carson-Jurica MA, Scharader WT, O’Malley BW: Steroid receptor family: structure and functions. Endocr Rev, 11: 201-222. 1990.
- Kimura N, Mizokami A, Oonuma T, Sasano H, Nagura H: Immunocytochemical localization of androgen receptor with polyclonal antibody in paraffin-embedded human tissues. J Histochem Cytochem, 41: 671-678, 1993.
- Pelletier G, Labrie C, Labrie F: Localization of oestrogen receptor alpha, oestrogen receptor beta and androgen receptors in the rat reproductive organs. J Endocrinol, 165: 359-370, 2000.
- Liao S, Kokontis J, Sai T, Hiipakka RA: Androgen receptors: structures, mutations, antibodies and cellular dynamics. J Steroid Biochem, 34: 41-51, 1989.
- Chang CS, Kokontis J, Liao ST: Structural analysis of complementary DNA and amino acid sequences of human and rat androgen receptors. Proc Natl Acad Sci, 85: 7211-7215, 1988.
- Lubahn DB, Joseph DR, Sar M, Tan J, Higgs HN, Larson RE, French FS, Wilson EM: The human androgen receptor: complementary deoxyribonucleic acid cloning, sequence analysis and gene expression in prostate. Mol Endocrinol, 2: 1265-1275, 1988.
- Shan LX, Rodriguez MC, Janne OA: Regulation of androgen receptor protein and mRNA concentrations by androgens in rat ventral prostate and seminal vesicles and in human hepatoma cells. Mol Endocrinol, 4: 1636-1646, 1990.
- Zhu LJ, Hardy MP, Inigo IV, Huhtaniemi I, Bardin W, Moo-Young AJ: Effects of androgen on androgen receptor expression in rat testicular and epididymal cells: a quantitative immunohistochemical study. Biol Reprod, 63: 368-376, 2000.
- Sanborn BM, Caston LA, Chang C, Liao S, Speller R, Porter LD, Ku CY: Regulation of androgen receptor mRNA in rat Sertoli and peritubular cells. Biol Reprod, 45: 634-641, 1991.
- Blok LJ, Themmen APN, Peter AHFM, Trapman J, Baarends WM, Hoogerbrugge JW, Grootegoed JA: Transcriptional regulation of androgen receptor gene expression in Sertoli cells and other cells types. Mol Cell Endocrinol, 88: 153-164, 1992.
- Shan LX, Bardin CW, Ardí MP: Immunohistochemical analysis of androgen effects on androgen receptor expression in developing Leydig and Sertoli cells. Endocrinology, 138: 1259-1266, 1997.
- Cardone A, Angelini F, Varriale B: Autoregulation of estrogen and androgen receptor mRNAs and downregulation of androgen receptorr mRNA by estrogen in primary cultures of lizard testis cells. Gen Comp Endocrinol, 110: 227-236, 1998.
- Goyal HO, Bartol FF, Wiley AA, Khalil MK, Williams CS, Vig MM: Regulation of androgen and estrogen receptors in male excurrent ducts of the goat: an immunohistochemical study. Anat Rec, 250: 164-171, 1998.
- Desjardins C, Lopez MJ: Environmental cues evoke differential responses in pituitary-testicular function in deer mice. Endocrinol, 112: 1398-1406, 1983.
- Nelson RJ, Kita M, Blom JMC, Rhyne-Grey J: Photoperiod influences the critical caloric intake necessary to maintain reproduction among male deer mice (Peromyscus maniculatus). Biol. Reprod, 46: 226-232, 1992.
- Young KA, Zirkin BR, Nelson RJ: Testicular regression in response to food restriction and short photoperiod in White-Footed Mice (Peromyscus leucopus) is mediated by apoptosis. Biol Reprod, 62: 347-354, 2000.
- Bayne K: Revised guide for the care and use of laboratory animals available. Am Phys Soc Physiol, 39: 208-211, 1996.
- Pine AP, Jessop NS, Oldham JD: Maternal protein reserves and their influence on lactational performance in rats: the effects of dietary protein restriction and stage of lactation on milk composition. Br J Nutr, 72: 815-830, 1994.
- Ramos CF, Teixeira CV, Passos MCF, Pazos-Moura CC, Lisboa PC, Curty FH, Moura EG: Low-protein diet changes thyroid function in lactating rats. PSEBM, 224: 256-263, 2000.
- Reeves PG, Nielsen FH, Fahey GC: AIN-93 Purified diets for laboratory rodents: final report of the American Institute of Nutrition Ad Hoc Writing Committee on the reformulation of the AIN-76 rodent diet. J Nut, 123: 1939-1951, 1993.
- Fischbeck KL, Rasmussen KM: Effect of repeated reproductive cycles on maternal nutritional status, lactational performance and litter growth in ad libitum fed and chronically food-restricted rats. J Nutr, 117: 1967-1975, 1987.
- Yagil R, Etizion Z, Berlyne GM: Changes in rat milk quantity and quality due to variations in litter size and high ambient temperature. Lab Natl Sci, 26: 33-37, 1976.
- Bradford MM: A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem, 72: 248-254, 1976.
- Zar JH: Bioestatistical Analysis. New Jersey, Prentice-Hall, 1998.
- Nastiuk KL, Clayton DF: Seasonal and tissue-specific regulation of canary androgen receptor menssenger ribonucleic acid. Endocrinol, 134: 640-649, 1994.
- Gonzalez-Cadavid NF, Vernet D, Navarro AF, Rodriguez AJ, Swerdloff RS, Rajfer J: Up-regulation of the levels of androgen receptor and its mRNA by androgens in smooth-muscle cells from rat penis. Mol Cell Endocrinol, 90: 219-229, 1993.
- Console GM, Jurado SB, Oyhenart E, Ferese C, Pucciarelli H, Gomez Dumm CL: Morphometric and ultrastructural analysis of different pituitary cell populations in undernourished monkeys. Braz J Med Biol Res, 34: 65-74, 2001.
______________________
Received: August 16, 2001
Accepted after revision: October 6, 2001
_______________________
Correspondence address:
Dra. Cristiane da Fonte Ramos
Unidade de Pesquisa Urogenital
Av. 28 de Setembro, No. 87, fundos
Prédio da FCM - térreo
Rio de Janeiro, RJ, 20551-030, Brazil
Fax: + + (55) (21) 2587-6121
E-mail: cramos@uerj.br